


































Skeletal muscle is the biological motor; contractile proteins use ATP to generate force and shorten the muscle, so converting chemical energy into the mechanical work needed to make things move. Muscle output is transmitted to the environment through elastic tendons, which connect muscle to a rigid skeleton, and via a gearing system created by the arrangement of linked elements of the skeleton. Such musculoskeletal systems produce the dazzling array of movements seen in animals as small as mites or as large as the blue whale; as fast as a cheetah or as slow as a sloth; and as strong as an ant or as delicate as a hummingbird.
Understanding the constraints to muscle performance is therefore fundamental to understanding biological movement and the staggering diversity in musculoskeletal system “design”. For example, larger vertebrate animals tend to have stouter bones and adopt a more upright posture, an adaptation thought to deal with the increasing demands of gravity. And elastic tendons that can recoil rapidly are thought to resolve the fundamental limitation shared by muscle and engineered motors: the force they generate typically declines at large velocities. This study combined the expertise in muscle physiology and elastic tissues of the HFSP Awardee Natalie C. Holt with that of the theoretical mechanical analyses of the HFSP Awardee David Labonte to examine how the molecular basis of force and work generation in muscle imposes elemental physical constraints on animal movement, and then used this novel understanding to explore the demands on musculoskeletal “design” across animal body sizes.
Consider jumping, a tractable and well-studied movement, as an example: the muscle provides the mechanical energy required to increase the body’s gravitational potential energy. The potential energy increase thus cannot exceed the work muscle can do - but how much work can muscle do? Historically, it has been assumed that each gram of muscle can do the same amount of work. This clearly makes some sense: a heavier motor should deliver more energy than a small motor, and probably roughly by as much as it is heavier. However, the problem with this perspective is that it ignores the dimension of time: muscle work cannot be delivered arbitrarily fast, and so the energy delivered may be limited by the rate at which it is delivered or the muscle’s power. But how much time do animals have to deliver muscle work? It turns out there is no unique answer to this question, and a muscle operating with fixed power can achieve a range of jump performances provided it remains below the universal limit posed by its work capacity. So, if it is neither the muscle’s power nor work capacity that limits jump performance in these instances, then what is?
Combining the conservation of energy with fundamental physiological aspects of muscle work and power production, the HFSP Research Grant awardees Holt and Labonte explore alternative constraints on muscle mechanical output. They show that the energy a muscle can deliver is limited not only by the distance through which it can shorten - the total work that it can do - but also by the peak velocity with which it can shorten. To gain an intuitive understanding of this limit, imagine yourself on a bike, cycling down a hill in a low gear. Although you would have no problem providing the force needed to move your legs, you may find yourself unable to accelerate the bike further because you simply cannot pedal fast enough – you are limited by how quickly your muscles can shorten, not by the force they generate. While, like all analogies, this picture is overly simplistic, it provides a sense of how a speed limit can prevent a strong motor like muscle from delivering its maximum work. Therefore, in addition to its work capacity, muscle energy output is also limited by what we termed its kinetic energy capacity.
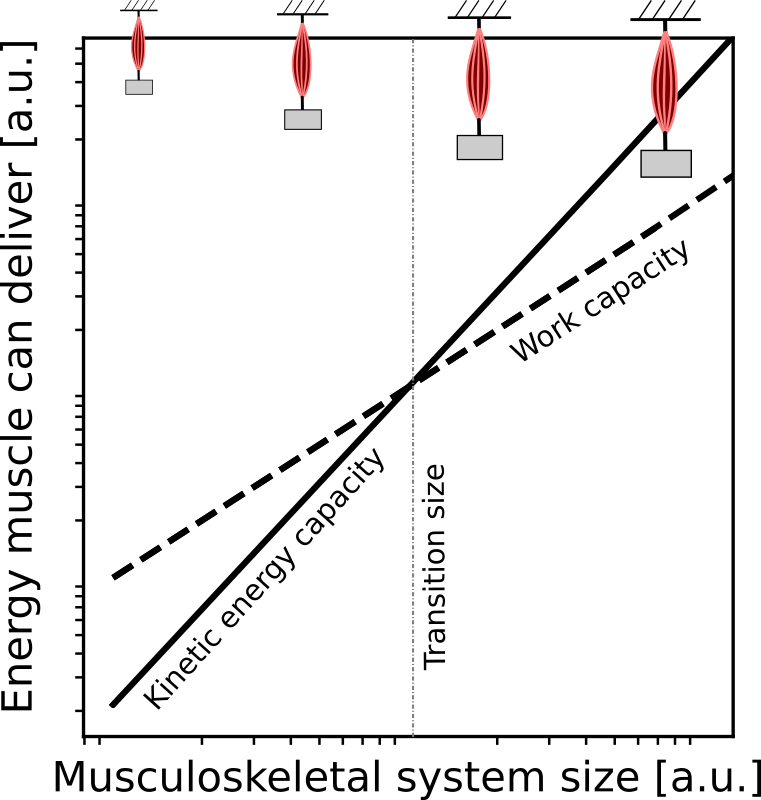
The kinetic energy capacity shares many features with the historical idea of a power limit and may thus appear only superficially different. However, a closer look reveals several important, if sometimes subtle, differences. Firstly, the kinetic energy capacity provides a unique limit to muscle performance, unlike the power capacity. Secondly, in contrast to the consistent maximum work and power output per unit muscle mass, the maximum kinetic energy capacity per muscle mass does change with muscle size. A rough estimate suggests that terrestrial animals up to ~50kg are limited by their kinetic energy capacity, whereas larger animals are limited by their work capacity. This would provide a tentative explanation for several well-established but not satisfactorily explained phenomena in comparative biomechanics, such as why the fastest animals are of intermediate size or why larger animals tend to adopt a more upright posture. Lastly, a considerable body of work has been dedicated to the amplification of muscle power by the recoil of elastic tendons. While tendons certainly amplify power, these findings suggest that this is an epiphenomenon and that the benefit may, in fact, be the amplification of the muscle’s kinetic energy capacity. Elastic recoil may be an adaptation that works around the fundamental physical constraints on muscle energy output that are particularly acute for animals with small body sizes.